Una Serie de Ojos
¿Cómo vemos? En el siglo XIX, la anatomía del ojo era conocida con gran detalle y los mecanismos sofisticados que utiliza para entregar una representación precisa del mundo exterior causaban asombro en cualquier persona que los conociera. Los científicos del siglo XIX observaron correctamente que si una persona tuviera la desdicha de perder uno de los muchos rasgos integrados del ojo, como ser la lente, o el iris, o los músculos oculares, el resultado inevitable sería una pérdida seria de visión o la ceguera definitiva. En consecuencia, se llegó a la conclusión que el ojo sólo podía funcionar si estaba prácticamente intacto.
Mientras Charles Darwin consideraba las posibles objeciones a su teoría de la evolución mediante la selección natural en El Origen de las Especies, abordó el problema del ojo en una sección del libro titulado apropiadamente "Órganos de Perfección y Complicación Extrema". Se dio cuenta que si aparecía en una generación un órgano de la complejidad del ojo, el evento sería sinónimo de un milagro. De alguna forma, para que la evolución darwiniana fuera creíble, debía eliminarse la dificultad que tenía el público para visualizar la formación gradual de órganos complejos.
Darwin tuvo un éxito brillante, no describiendo efectivamente un camino que la evolución podría haber tomado al construir el ojo, sino más bien señalando una cantidad de animales que se sabía que tenían ojos de distintas estructuras, pasando desde el simple punto sensible a la luz hasta el ojo complejo de los vertebrados, similar a la cámara fotográfica, y sugiriendo que la evolución del ojo humano pudo haber involucrado órganos similares como intermediarios.
Pero la pregunta persiste: ¿cómo vemos? Si bien Darwin logró persuadir a gran parte del mundo de que el ojo moderno podía ser producido gradualmente desde una estructura mucho más simple, ni siquiera intentó explicar cómo el simple punto sensible a la luz, que era su punto de partida, funcionaba realmente. Al discutir el ojo, Darwin descartó el asunto de su mecanismo último:{1}
Cómo un nervio llega a ser sensible a la luz difícilmente nos interese más que el tema de cómo se originó la vida misma.
Él tenía una excelente razón para declinar contestar la pregunta: la ciencia del siglo XIX no había progresado al punto en que el tema pudiera ser abordado siquiera. El asunto de cómo funciona el ojo - es decir, qué ocurre cuando el fotón de luz primero impacta a la retina - simplemente no podía ser contestado en ese tiempo. A decir verdad, ninguna cuestión acerca del mecanismo subyacente de la vida podía ser contestada en ese tiempo. ¿Cómo provocan el movimiento los músculos de los animales? ¿Cómo funciona la fotosíntesis? ¿Cómo es extraída la energía de los alimentos? ¿Cómo combate el cuerpo la infección? Nadie lo sabía.
El Calvinismo
Ahora bien, parece ser una característica de la mente humana que, cuando no está limitada por el conocimiento de los mecanismos de un proceso, le parezca fácil imaginarse los pasos que van de la no-función a la función. Un buen ejemplo de esto puede verse en la tira cómica popular, Calvin y Hobbes. El niñito Calvin siempre está teniendo aventuras acompañado por su tigre Hobbes, saltando dentro de una caja y viajando hacia atrás en el tiempo, o tomando una pistola de rayos de juguete y "transmogrifándose" en distintas formas animales, o nuevamente usando la caja como una duplicadora y haciendo copias de sí mismo para poder enfrentar poderes terrenales, como su mamá y sus maestras. A un pequeño niño como Calvin le resulta fácil imaginarse que una caja pueda volar como un avión (o algo), porque Calvin no sabe cómo funcionan los aviones.
Un buen ejemplo tomado del mundo biológico de cambios complejos que parecen simples es la creencia en la generación espontánea. Uno de los principales proponentes de la teoría de la generación espontánea a mediados del siglo XIX fue Ernst Haeckel, un gran admirador de Darwin y un difusor de la teoría de Darwin. Desde la visión limitada de las células que proveían los microscopios del siglo XIX, Haeckel creía que una célula era un "simple pedacito de una combinación albuminosa de carbono",{2} no muy diferente de una porción microscópica de gelatina. En consecuencia, le parecía a Haeckel que una vida de este tipo podría ser producida fácilmente desde la materia inanimada. En 1859, el año de la publicación de El Origen de las Especies, un barco exploratorio, el H.M.S. Cyclops, excavó un tipo de barro de apariencia extraña del fondo del mar. Eventualmente Haeckel llegó a observar el barro y pensó que se parecía mucho a unas células que había visto bajo el microscopio. Se lo mostró excitadamente a Thomas Henry Huxley, el gran amigo y defensor de Darwin. Huxley también se convenció que era Urschleim (es decir, protoplasma), el progenitor de la vida misma, y Huxley llamó al barro Bathybius Haeckelii, en honor al eminente proponente de la abiogénesis.
El barro no llegó a crecer. En años posteriores, con el desarrollo de nuevas técnicas bioquímicas y microscopios mejorados, la complejidad de la célula fue revelada. Se demostró que los "simples pedacitos" contenían miles de diferentes tipos de moléculas orgánicas, proteínas, ácidos nucleicos, muchas estructuras discretas subcelulares, compartimentos especializados para procesos especializados, y una arquitectura extremadamente compleja. Mirando hacia atrás desde la perspectiva de nuestro tiempo, el episodio del Bathybius Haeckelii parece tonto o completamente embarazoso, pero no debería serlo. Haeckel y Huxley estaban comportándose naturalmente, como Calvin: como no estaban al tanto de la complejidad de las células, encontraron que era fácil creer que las células podían originarse del simple barro.
A lo largo de la historia ha habido muchos ejemplos, similares al de Haeckel, Huxley y la célula, donde una pieza clave de un rompecabezas científico en particular estaba más allá de la comprensión de la época. En la ciencia hasta existe un término caprichoso para una máquina o estructura o proceso que hace algo, pero cuyo mecanismo para lograrlo es desconocido: se lo llama "caja negra." En el tiempo de Darwin toda la biología era una caja negra: no sólo la célula, o el ojo, o la digestión, o la inmunización, sino cada estructura y función biológica porque, en última instancia, nadie podía explicar cómo ocurrían los procesos biológicos.
Ernst Mayr, el destacado biólogo, historiador, y fuerza guía detrás de la síntesis neo-darwiniana, ha señalado que:{3}
Toda revolución científica necesita aceptar toda suerte de cajas negras, porque si uno tuviera que esperar hasta que fueran abiertas todas las cajas negras uno nunca tendría progresos conceptuales.
Eso es cierto. Pero en el pasado, cuando las cajas negras fueron abiertas finalmente, la ciencia, y a veces el mundo entero, pareció cambiar. La biología ha progresado tremendamente debido al modelo que postuló Darwin. Pero las cajas negras que aceptaba Darwin han sido abiertas ahora, y nuestra visión del mundo está siendo sacudida nuevamente.
Las Proteínas
A fin de entender la base molecular de la vida es necesario entender cómo funcionan unas cosas llamadas "proteínas." Aunque la mayoría de la gente piensa en las proteínas como "algo que se come", uno de los principales grupos de alimentos, cuando están dentro de un animal o una planta que no ha sido comido las proteínas cumplen un propósito diferente. Las proteínas son la maquinaria del tejido vivo que construye las estructuras y lleva a cabo las reacciones químicas necesarias para la vida. Por ejemplo, el primero de los muchos pasos necesarios para la conversión del azúcar a formas biológicamente utilizables de energía es llevado a cabo por una proteína llamada hexoquinasa. La piel está formada, en gran medida, por una proteína llamada colágeno. Cuando la luz impacta sobre su retina, interactúa primeramente con una proteína llamada rhodopsina. Como puede verse aun por este número limitado de ejemplos, las proteínas llevan a cabo funciones asombrosamente diversas. Sin embargo, en general una proteína dada puede llevar a cabo sólo una o unas pocas funciones: la rhodopsina no puede formar piel, y el colágeno no puede interactuar en forma útil con la luz. De ahí que una célula típica contenga miles y miles de distintos tipos de proteínas que realizan las muchas tareas necesarias para la vida, en forma muy parecida al taller de un carpintero que podría contener muchos tipos diferentes de herramientas para distintos trabajos de carpintería.
¿A qué se parecen estas herramientas versátiles? La estructura básica de las proteínas es bastante sencilla: se forman enlazando en una cadena sub-unidades discretas llamadas aminoácidos. Si bien la cadena de la proteína puede estar formada por una cantidad de enlaces de aminoácidos que puede ir desde 50 a 1000, cada posición sólo puede contener uno entre veinte aminoácidos diferentes. En este sentido son muy parecidas a las palabras: las palabras vienen en distintos largos, pero están formadas por un conjunto discreto de 26 letras (en inglés). Ahora bien, una proteína en una célula no está flotando como una cadena fláccida; más bien, se enrolla en una estructura muy precisa que puede ser bastante diferente para distintos tipos de proteínas. Al fin y al cabo, dos secuencias diferentes de aminoácidos - dos proteínas diferentes - pueden ser enrolladas en estructuras tan específicas y diferentes entre sí como una llave de tres octavos de pulgada puede ser diferente de una sierra. Y, al igual que las herramientas caseras, si la estructura de las proteínas está deformada en forma significativa, entonces ellas no logran realizar sus cometidos.
La Visión Humana
En general, los procesos biológicos a nivel molecular son realizados por redes de proteínas, cada uno de cuyos integrantes lleva a cabo una tarea en particular en una cadena.
Volvamos a la pregunta de cómo vemos. Aunque para Darwin el evento principal de la visión era una caja negra, gracias a los esfuerzos de muchos bioquímicos tenemos a nuestra disposición una respuesta a la pregunta de la visión.{4} Cuando la luz llega a la retina, un fotón es absorbido por una molécula orgánica llamada 11-cis-retinal, lo que hace que se reacomode en unos picosegundos en trans-retinal. El cambio de forma del retinal provoca un cambio de forma correspondiente en la proteína, rhodopsina, a la que está fuertemente ligada. Como consecuencia de la metamorfosis de la proteína, el comportamiento de la proteína cambia en una forma muy específica. La proteína alterada puede interactuar ahora con otra proteína llamada transducina. Antes de asociarse con la rhodopsina, la transducina está fuertemente ligada a una pequeña molécula orgánica llamada GDP, pero cuando se liga a la rhodopsina la GDP de disocia de la transducina, y una molécula llamada GTP, muy relacionada pero críticamente diferente del GDP, se liga a la transducina.
El reemplazo de GTP por GDP en el complejo transducinarhodopsina altera su comportamiento. La GTP-transducinarhodopsina se liga a una proteína llamada fosfodiesterasa, ubicada en la membrana interna de la célula. Cuando está ligada por la rhodopsina y su séquito, la fosfodiesterasa adquiere la habilidad de hendir químicamente una molécula llamada cGMP. Inicialmente, hay muchas moléculas cGMP en la célula, pero la acción de la fosfodiesterasa disminuye la concentración de cGMP. Activar la fosfodiesterasa puede compararse con sacar el tapón de una bañadera, haciendo que disminuya el nivel del agua.
Una segunda proteína-membrana que se liga a la cGMP, llamada canal iónico, puede pensarse como una compuerta especial que regula la cantidad de iones de sodio en la célula. El canal iónico normalmente permite que los iones de sodio fluyan hacia dentro de la célula, mientras que otra proteína aparte los bombea activamente hacia fuera de nuevo. La acción dual del canal iónico y de las proteínas bombeadoras mantiene el nivel de los iones de sodio en la célula dentro de un rango estrecho. Cuando la concentración de cGMP se reduce respecto de su valor normal mediante la hendidura de la fosfodiesterasa, muchos canales se cierran, lo que resulta en una concentración celular disminuida de iones de sodio con carga positiva. Esto provoca un desequilibrio de cargas a ambos lados de la membrana de la célula que, finalmente, hace que una corriente sea transmitida por el nervio óptico al cerebro. El resultado, cuando es interpretado por el cerebro, es la visión.
Si la bioquímica de la visión estuviera limitada a las reacciones detalladas más arriba, la célula rápidamente se vaciaría de su aprovisionamiento de 11-cis-retinal y de cGMP, a la vez que se vaciaría de iones de sodio. Por lo tanto, se requiere un sistema que limite la señal que se genera y que restaure la célula a su estado original. Hay varios mecanismos que hacen esto. Normalmente, en la oscuridad, el canal iónico, además de iones de sodio, también permite que entren iones de calcio a la célula. El calcio es bombeado hacia fuera por una proteína diferente a fin de mantener una concentración intracelular de calcio constante. Sin embargo, cuando los niveles de cGMP caen, cerrando el canal de iones y disminuyendo la concentración de iones de sodio, la concentración de iones de calcio es reducida también. La enzima fosfodiesterasa, que destruye a la cGMP, es frenada considerablemente ante una concentración de calcio menor. Además, una proteína llamada guanilata ciclasa comienza a resintetizar la cGMP cuando los niveles de calcio comienzan a disminuir. Entretanto, mientras todo esto está ocurriendo, la metarhodopsina II es modificada químicamente por una enzima llamada rhodopsina quinasa, que coloca un grupo fosfato en su sustrato. La rhodopsina modificada es ligada entonces por una proteína llamada arrestina, que impide que la rhodopsina siga activando a la transducina. En consecuencia, la célula contiene mecanismos que limitan la señal amplificada activada por un solo fotón.
El trans-retinal eventualmente se sale de la molécula de rhodopsina y debe ser reconvertida a 11-cis-retinal y ligada nuevamente por la opsina para regenerar rhodopsina para otro ciclo visual. Para lograr esto, primero el trans-retinal es modificado químicamente por una enzima a transretinol, una forma que contiene dos átomos de hidrógeno más. Una segunda enzima entonces isomeriza la molécula de 11-cis-retinol. Finalmente, una tercera enzima quita los átomos de hidrógeno agregados anteriormente para formar 11-cis-retinal, y el ciclo está completo.
Explicar la Vida
Aunque muchos de los detalles de la bioquímica de la visión no han sido mencionados aquí, el resumen que acabamos de ver tiene la intención de demostrar que, en última instancia, esto es lo que significa "explicar" la visión. Éste es el nivel de explicación al que debe apuntar la ciencia biológica finalmente. Para decir que uno entiende alguna función, cada paso relevante del proceso deber ser explicado. Los pasos relevantes en los procesos biológicos ocurren en última instancia en el nivel molecular, así que una explicación satisfactoria de un fenómeno biológico como la visión, o la digestión, o la inmunización, debe incluir una explicación molecular. Ya no es suficiente, ahora que la caja negra de la visión ha sido abierta, que una "explicación evolucionista" invoque solamente las estructuras anatómicas de los ojos, como lo hizo Darwin en el siglo XIX y como lo siguen haciendo la mayoría de los difusores de la evolución hoy día. La anatomía es, simplemente, irrelevante. También lo es el registro fósil. No importa si el registro fósil es consistente o no con la teoría evolucionista, así como no importó en la física que la teoría de Newton fuera consistente con la experiencia cotidiana. El registro fósil no tiene nada que decirnos acerca de, digamos, si las interacciones del 11-cis-retinal con la rhodopsina, la transducina y la fosfodiesterasa se podrían haber desarrollado paso a paso. Tampoco importan los patrones de la biogeografía, o la genética de las poblaciones, o las explicaciones que la teoría evolucionista ha dado para los órganos rudimentarios o la abundancia de las especies.
"Cómo un nervio llega a ser sensible a la luz difícilmente nos interese más que el tema de cómo se originó la vida misma", dijo Darwin en el siglo XIX. Pero ambos fenómenos han atraído el interés de la bioquímica moderna. La historia del lento parálisis de la investigación sobre el origen de la vida es bastante interesante, pero el espacio nos impide volver a contarla aquí. Es suficiente decir que al presente el campo de los estudios sobre el origen de la vida se ha disuelto en una cacofonía de modelos contradictorios, cada uno de ellos poco convincente, seriamente incompleto, e incompatible con los modelos competidores. En privado, aun los biólogos más evolucionistas admitirán que la ciencia no tiene ninguna explicación para el origen de la vida.{5}
El propósito de este ensayo es mostrar que los mismos problemas que asedian a la investigación sobre el origen de la vida también hostigan los esfuerzos por mostrar cómo llegó a existir prácticamente cualquier sistema bioquímico complejo. La bioquímica ha revelado un mundo molecular que resiste decididamente la explicación por la misma teoría que ha sido aplicada desde tiempo atrás al nivel de todo el organismo. Ninguna de las cajas negras de Darwin - el origen de la vida, o el origen de la visión, u otros sistemas bioquímicos complejos - ha sido explicada por su teoría.
Complejidad Irreducible
En El Origen de las Especies Darwin declaró:{6}
Si pudiera demostrarse que existe algún órgano complejo que no pudiera haberse formado, de alguna forma, a partir de modificaciones numerosas, sucesivas y leves, mi teoría se vendría abajo completamente.
Un sistema que cumple con el criterio de Darwin es aquél que exhibe complejidad irreducible. Por complejidad irreducible quiero decir un único sistema que está compuesto por varias partes interactuantes que contribuyen a la función básica, y en el que la remoción de cualquiera de las partes hace que el sistema deje de funcionar efectivamente. Un sistema irreduciblemente complejo no puede ser formado gradualmente por modificaciones leves y sucesivas de un sistema precursor, ya que cualquier precursor a un sistema irreduciblemente complejo es, por definición, no-funcional. Dado que la selección natural necesita una función para seleccionar, un sistema biológico irreduciblemente complejo, si existiera tal cosa, tendría que surgir como una unidad integrada para que la selección natural pudiera tener algo sobre lo cual actuar. Está aceptado casi universalmente que un evento abrupto de este tipo sería irreconciliable con el gradualismo que visualizaba Darwin. En este punto, sin embargo, "irreduciblemente complejo" es sólo un término, cuyo poder reside mayormente en su definición. Debemos preguntarnos ahora si existe alguna cosa real que sea de hecho irreduciblemente compleja y, si es así, si hay entonces cosas irreduciblemente complejas que sean sistemas biológicos también.
Considere la humilde trampa para cazar ratones (Figura 1). Las trampas para ratones que usa mi familia en nuestro hogar para tratar con roedores molestos consisten de varias partes. Estas son: (1) una plataforma de madera plana que actúa como una base; (2) un martillo de metal que realiza la verdadera tarea de aplastar al ratoncito; (3) un resorte de alambre con extremos extendidos para hacer presión contra la plataforma y el martillo cuando se carga la trampa; (4) una traba sensible que se suelta cuando se le aplica una leve presión; y (5) una barra de retención que retiene al martillo cuando se carga la trampa y que se conecta con la traba. Hay también una cantidad variada de grapas y tornillos que mantienen unido al sistema.
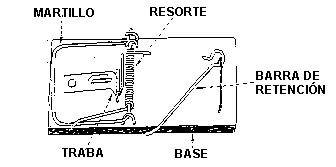
Figura 1. Una trampa para cazar ratones casera. Las partes funcionales de la trampa están rotuladas. Si alguna de las partes falta, la trampa no funciona.
Si cualquiera de los componentes de la trampa (la base, el martillo, el resorte, la traba o la barra) es eliminado, entonces la trampa no funciona. En otras palabras, la simple trampa para cazar ratones no tiene la capacidad de atrapar al ratón hasta que varias partes separadas estén todas ensambladas.
Dado que la trampa está compuesta necesariamente de varias partes, es irreduciblemente compleja. En consecuencia, existen los sistemas irreduciblemente complejos.
Máquinas Moleculares
Ahora bien, ¿hay sistemas bioquímicos que sean irreduciblemente complejos? Sí, resulta que muchos de ellos lo son.
Anteriormente discutimos las proteínas. En muchas estructuras biológicas las proteínas son simplemente componentes de máquinas moleculares más grandes. Como el tubo de rayos catódicos, los cables, las tuercas y los tornillos que forman el televisor, muchas proteínas son parte de estructuras que sólo funcionan cuando prácticamente todos los componentes han sido ensamblados. Un buen ejemplo de esto es la cilia.{7}
 Figura 2a. Animación de una Cilia
Figura 2a. Animación de una Cilia
Las cilias son organelas en forma de pelos que están en las superficies de muchas células de los animales y de las plantas inferiores, y que sirven para mover los fluidos sobre la superficie de la célula, o como "remos" para que las células individuales se puedan mover a través de un fluido. En los humanos, por ejemplo, cada una de las células epiteliales que recubren el tracto respiratorio tiene unas 200 cilias que se mueven en forma sincronizada para mover el moco hacia la garganta para que sea eliminado. Una cilia consiste de un manojo de fibras cubierto por una membrana denominado axonema. Un axonema contiene un anillo de 9 microtubos dobles que rodean 2 microtubos simples centrales. Cada par externo consiste de un anillo de 13 filamentos (subfibra A) fundido con un conjunto de 10 filamentos (subfibra B). Los filamentos de los microtubos están formados por dos proteínas llamadas alfa y beta tubulina. Los 11 microtubos que forman un axonema se mantienen unidos por tres tipos de conectores: las subfibras A están unidas a los microtubos centrales por rayos radiales; los pares adyacentes externos están unidos por uniones que consisten de una proteína muy elástica llamada nexina; y los microtubos centrales están unidos por un puente conector. Finalmente, cada subfibra A tiene dos brazos, un brazo interno y un brazo externo, cada uno de los cuales contiene la proteína dineína.
Pero, ¿cómo funciona la cilia? Los experimentos han indicado que el movimiento ciliar resulta del "caminar" accionado químicamente de los brazos de dineína en un microtubo sobre la subfibra B vecina de un segundo microtubo, de forma que los dos microtubos se deslizan uno a lo largo del otro (Figura 2a y b). Sin embargo, las uniones cruzadas de proteínas entre los microtubos en una cilia intacta impiden que los microtubos vecinos se deslicen uno sobre el otro más que una corta distancia. Estas uniones cruzadas, por lo tanto, convierten al movimiento deslizante inducido por la dineína en un movimiento de curvado de todo el axonema.
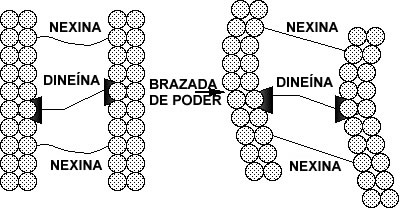
Figura 2b. Dibujo esquemático de parte de una cilia. La brazada de poder de la proteína motor, dineína, fijada a un microtubo, contra la subfibra B de un microtubo vecino hace que la fibras se deslicen una sobre otra. La proteína conectora flexible, nexina, convierte al movimiento deslizante en un movimiento de curvado.
Ahora, detengámonos un poco, y revisemos el funcionamiento de la cilia, y consideremos lo que implica. Las cilias están compuestas de por lo menos una media docena de proteínas: alfa-tubulina, beta-tubulina, dineína, nexina, la proteína-rayo, y una proteína-puente central. Estas se combinan para realizar una tarea, el movimiento ciliar, y todas estas proteínas deben estar presentes para que funcione la cilia. Si las tubulinas están ausentes, no hay filamentos que puedan deslizarse; si la dineína está ausente, entonces la cilia se mantiene rígida y quieta; si la nexina u otras proteínas conectoras están ausentes, entonces el axonema se rompe cuando los filamentos se deslizan.
Lo que vemos en la cilia, entonces, no es sólo complejidad profunda, sino complejidad irreducible en la escala molecular. Recuerde que al decir "complejidad irreducible" queremos decir un aparato que requiere varios componentes distintos para que el todo pueda funcionar. Mi trampa debe tener una base, un martillo, un resorte, una traba, una barra, todos trabajando en conjunto, para que pueda funcionar. De la misma forma, la cilia, tal como está constituida, debe tener los filamentos deslizantes, las proteínas conectoras, y las proteínas motoras para que ocurra la función. Si falta alguno de esos componentes el aparato es inservible.
Los componentes de una cilia son moléculas simples. Esto significa que ya no hay más cajas negras para invocar; la complejidad de la cilia es final, fundamental. Y, de la misma forma que los científicos, cuando empezaron a comprender la complejidades de la célula, se dieron cuenta de cuán tonto era pensar que la vida surgió en forma espontánea en un solo paso o varios pasos del barro del océano, así también nosotros ahora nos damos cuenta que la cilia compleja no puede conseguirse en un solo paso o en unos pocos pasos. Pero, como la complejidad de la cilia es irreducible, entonces no puede tener precursores funcionales. Como la cilia irreduciblemente compleja no puede tener precursores funcionales no puede ser producida por la selección natural, la que requiere de un continuo de funciones para operar. La selección natural es impotente cuando no hay ninguna función para seleccionar. Podemos ir más lejos y decir que, si la cilia no puede ser producida por selección natural, entonces la cilia fue diseñada.
El Estudio de la "Evolución Molecular"
Abundan otros ejemplos de complejidad irreducible, incluyendo aspectos del transporte de proteínas, la coagulación de la sangre, el ADN circular cerrado, el flagelo bacterial, los telómeros, la fotosíntesis, la regulación de la transcripción y muchos más. Se pueden encontrar ejemplos de complejidad irreducible en prácticamente cada página de un libro de texto de bioquímica. Pero si estas cosas no pueden explicarse por la evolución darwiniana, ¿cómo ha considerado estos fenómenos la comunidad científica durante los últimos cuarenta años? Un buen lugar para contestar esa pregunta es el Journal of Molecular Evolution (Revista de la Evolución Molecular). El JME es una revista especializada que fue iniciada específicamente para tratar con el tema de cómo ocurre la evolución a nivel molecular. Tiene altos estándares científicos y es editado por figuras destacadas en el campo. En un número reciente de JME se publicaron once artículos; de estos, la totalidad de los once estaban dedicados al análisis de las secuencias de las proteínas y del ADN. Ninguno de estos ensayos trataban modelos detallados para los intermediarios en el desarrollo de estructuras biomoleculares complejas. En los últimos diez años JME ha publicado 886 artículos. De estos, 95 trataban la síntesis química de moléculas consideradas necesarias para el origen de la vida, 44 proponían modelo matemáticos para mejorar el análisis de las secuencias, 20 trataban con las implicaciones evolucionistas de las estructuras actuales y 719 eran análisis de las secuencias de proteínas o de polinucleótidos. Hubo una cantidad cero de artículos que trataran modelos detallados para los intermediarios en el desarrollo de estructuras biomoleculares complejas. Esta no es una peculiaridad de JME. No pueden encontrarse artículos que traten modelos detallados de los intermediarios en Proceedings of the National Academy of Science, Nature, Science, Journal of Molecular Biology o, que yo sepa, en ninguna revista especializada existente.
Las comparaciones de secuencias dominan en forma apabullante la literatura de la evolución molecular. Pero las comparaciones de secuencias simplemente no pueden explicar el desarrollo de sistemas biomoleculares complejos, de la misma forma que la comparación de Darwin entre ojos sencillos y complejos no le pudo decir cómo funcionaba la visión. En consecuencia, en esta área la ciencia está muda. Esto significa que cuando inferimos que los sistemas bioquímicos complejos fueron diseñados, no estamos contradiciendo ningún resultado experimental, no estamos en conflicto con ningún estudio teórico. No hace falta cuestionar ningún experimento, pero la interpretación de todos los experimentos deben ser reexaminados ahora, así como los experimentos que eran consistentes con un punto de vista newtoniano del universo tuvieron que ser reinterpretados cuando se descubrió la dualidad de la materia como onda y como partícula.
Conclusión
A menudo se dice que la ciencia debe evitar conclusiones que tienen un sabor a lo sobrenatural. Pero esto me parece a mí que es mala lógica y mala ciencia a la vez. La ciencia no es un juego donde se usan reglas arbitrarias para decidir cuáles explicaciones deben ser permitidas. Más bien, es un esfuerzo por hacer afirmaciones verdaderas acerca de la realidad física. Hace sólo sesenta años que el universo fue observado por primera vez. Este hecho sugirió inmediatamente un evento singular - que en algún momento en el pasado distante el universo comenzó a expandirse a partir de un tamaño extremadamente pequeño. Para mucha gente esta inferencia estaba cargada con alusiones a un evento sobrenatural - la creación, el comienzo del universo. El destacado físico A. S. Eddington tal vez habló por muchos físicos al expresar su disgusto con tal idea:{8}
Filosóficamente, la idea de un comienzo abrupto del orden presente de la Naturaleza me resulta repugnante, como considero que debe serlo para la mayoría; y aun aquellos a quienes les gustaría una prueba de la intervención de un Creador probablemente consideren que darle cuerda al universo una única vez en una época remota no es realmente el tipo de relación entre un Dios y su mundo que trae satisfacción a la mente.
No obstante, la hipótesis del Big Bang fue abrazada por los físicos y, con los años, ha mostrado ser un paradigma muy fructífero. El punto aquí es que la física siguió a los hechos adonde parecían conducir, aun cuando algunos pensaban que el modelo daba aliento y consuelo a la religión. En la actualidad, a medida que la bioquímica multiplica los ejemplos de sistemas moleculares fantásticamente complejos, sistemas que desalientan aun el intento de explicar cómo podrían haber surgido, debemos aprender de la física. La conclusión del diseño fluye en forma natural de los hechos; no debemos tenerle miedo; debemos abrazarla y construir a partir de ella.
En conclusión, es importante que nos demos cuenta que no estamos infiriendo el diseño a partir de lo que no sabemos, sino a partir de lo que sí sabemos. No estamos infiriendo el diseño para explicar una caja negra, sino para explicar una caja abierta. Un hombre de una cultura primitiva que ve un automóvil podría suponer que está accionado por el viento o por un antílope escondido bajo el automóvil, pero cuando abre la capota y ve el motor inmediatamente se da cuenta que estaba diseñado. De la misma forma, la bioquímica ha abierto la célula para examinar lo que lo hace funcionar y vemos que ella también ha sido diseñada.
Fue una conmoción para la gente del siglo diecinueve cuando descubrieron, a partir de las observaciones que la ciencia había hecho, que muchos rasgos del mundo biológico podían ser atribuidos al elegante principio de la selección natural. Es una conmoción para nosotros en el siglo veinte descubrir, a partir de observaciones que ha hecho la ciencia, que los mecanismos fundamentales de la vida no pueden ser atribuidos a la selección natural y, por lo tanto, fueron diseñados. Pero debemos ocuparnos de nuestra conmoción lo mejor que podamos y seguir. La teoría de la evolución no dirigida ya está muerta, pero el trabajo de la ciencia continúa.
Referencias
{1}Darwin, Charles (1872) Origin of Species, 6th ed (1988), p.151, New York University Press, New York. Volver al texto
{2}Farley, John (1979) The Spontaneous Generation Controversy from Descartes to Oparin, 2nd ed, p.73, The Johns Hopkins University Press, Baltimore. Volver al texto
{3}Mayr, Ernst (1991) One Long Argument, p. 146, Harvard University Press, Cambridge. Volver al texto
{4}Devlin, Thomas M. (1992) Textbook of Biochemistry, pp.938954, WileyLiss, New York. Volver al texto
{5}El retórico John Angus Campbell de la University of Washington ha notado que "los grandes edificios de ideas tales como el positivismo nunca mueren realmente. La gente pensante gradualmente los abandonan, pero se guardan las partes que son persuasivamente útiles para espantar a los que no están informados." "The Comic Frame and the Rhetoric of Science: Epistemology and Ethics in Darwin's Origin," Rhetoric Society Quarterly 24, pp.2750 (1994). Esto se aplica por cierto a la forma en que la comunidad científica maneja las cuestiones con relación al origen de la vida. Volver al texto
{6}Darwin, p.154. Volver al texto
{7}Voet, D. & Voet, J.G. (1990) Biochemistry, pp.11321139, John Wiley & Sons, New York. Volver al texto
{8}Citado en Jaki, Stanley L. (1980) Cosmos and Creator, pp.56, Gateway Editions, Chicago. Volver al texto
Copyright © 1997 Michael Behe. Todos los derechos reservados.
